gradiente electroquímico combina el concepto matemático de gradiente con los conceptos físicos y químicos de potencial eléctrico y de potencial químico (concentración). Básicamente indica cuál es la dirección en la que cambia más rápidamente la concentración y el potencial eléctrico de una solución no homogénea; esto es importante porque una partícula de una sustancia cualquiera con una cierta carga en solución se moverá tratando de seguir la dirección de mayor gradiente electroquímico, yendo desde donde esa sustancia en particular se encuentra más concentrada hacia donde está más diluida y desde donde tiene mayor potencial eléctrico hacia donde tiene menor potencial eléctrico.
Los gradientes electroquímicos son muy importantes para los seres vivos, donde son utilizados para provocar movimiento de sustancias a través de las membranas celulares y para producir energía aprovechable a través del proceso de fosforilación oxidativa.
Conceptualización[editar]
Gradiente[editar]
Un gradiente es, básicamente, una serie de vectores asociados a determinados puntos del espacio (campo vectorial), que indican cuáles son las direcciones de mayor cambio en un campo escalar (esto es una serie de puntos en el espacio con determinados valores escalares asociados).
Para entender el caso particular del potencial electroquímico se puede pensar en principio en un mapa de líneas de nivel de una zona geográfica determinada, este será nuestro "campo"; donde a cada coordenada espacial (latitud, longitud) se le asocia una determinada altura (valor escalar). el gradiente sería como una serie de flechas paralelas (tangentes en realidad) a la superficie que indicarían en que dirección se inclina más esa superficie.
Y aproximándonos aún más, podemos imaginarnos una habitación lo suficientemente grande (campo tridimensional), en la que hay un gas desigualmente distribuido. En las zonas donde hay más gas la presión (valor escalar) ejercida por el mismo es mayor que en las zonas donde hay menos gas. Aquí podemos imaginarnos el gradiente de presión como flechas que indican las direcciones en las cuales hay mayor diferencia de presión dentro del espacio tridimensional de la habitación.
Gradiente de concentración[editar]
Si arrojamos un pequeño cristal de azúcar dentro de un vaso que contiene agua, podemos ver que al principio el cristal parece no experimentar cambios, pero si esperamos un tiempo suficientemente largo, el cristal se disuelve completamente y se puede demostrar que las moléculas de azúcar se han distribuido uniformemente dentro del volumen del líquido. Entre la situación inicial y la final existe todo un proceso durante el cual las moléculas de azúcar se encuentran desigualmente distribuidas dentro del volumen del líquido. Cerca de la superficie del cristal hay una gran cantidad de moléculas de azúcar en relación a la cantidad de moléculas de agua (alta concentración); mientras que lejos de la superficie del cristal hay muy pocas moléculas de azúcar (baja concentración) entre ambos puntos existe todo un degradé de concentraciones. En este escenario existe un gradiente efectivo de concentración entre la zona cercana al cristal de azúcar y la zona más alejada. El concepto de gradiente es importante para comprender como es que a pesar de que el movimiento de cada una de las moléculas de azúcar dentro del líquido es aleatorio, existe un flujo neto de moléculas (difusión) desde la zona más cercana al cristal hacia las zonas más alejadas. Esto que es simplemente una consecuencia estadística de la suma de un gran número de movimientos aleatorios, se produce siguiendo las líneas de mayor diferencia de concentración, es decir de acuerdo al gradiente de concentración del azúcar dentro de la solución.
Generalidades[editar]
El potencial electroquímico es importante en la química electroanalítica y en la industria, en la cual tiene diversas aplicaciones (baterías, pilas de combustible…). Constituye una de las muchas maneras de intercambio de energía potencial en las cuales la energía se conserva.
En los procesos biológicos, la dirección de los movimientos iónicos realizados por difusión o transporte activo a través de una membrana está determinada por el gradiente electroquímico. En las mitocondrias y los cloroplastos, los gradientes de protones se utilizan para generar un potencial quimiosmótico (que es también conocido como fuerza motriz protónica). Esta energía potencial es utilizada para la síntesis de ATP a través de la fosforilación oxidativa.
Un gradiente electroquímico está constituido por dos componentes. En primer lugar, el componente eléctrico, el cual se origina debido a la diferencia de cargas a través de la membrana lipídica. En segundo lugar, el componente químico es debido a la diferencia de concentración de iones a ambos lados de la membrana. La combinación de estos dos factores determina la favorable dirección termodinámica para el movimiento de un ion a través de la membrana.
El gradiente electroquímico es análogo a la presión hidrostática a través de una presa hidroeléctrica. Proteínas de transporte de membrana, tales como la bomba de sodio-potasio, son equivalentes a las turbinas que convierten la energía potencial del agua en otras formas de energía física o química y los iones que atraviesan la membrana son equivalentes al agua que termina en la parte inferior de la presa. Además, la energía puede ser utilizada para bombear el agua hasta el lago que se encuentra encima de la presa. De modo similar, la energía química de las células puede ser utilizada para crear gradientes.
Química[editar]
El término se suele aplicar en contextos en los cuales se produce una reacción química, como los que implican la transferencia de un electrón en una batería de electrodos. En una batería, debido al movimiento de los iones, el potencial electroquímico crece y equilibra la energía de reacción de los electrodos. El voltaje máximo que se puede producir en una reacción de batería se llama a veces potencial electroquímico estándar de esa reacción (ver también potencial de electrodo y Tabla potenciales electroquímicos estándar). En los casos relacionados concretamente con el movimiento de solutos con carga eléctrica, el potencial es a menudo expresado en unidades de voltios. Ver: concentración celular.
Contexto biológico[editar]
En la biología, el término se usa a veces en el contexto de una reacción química, en concreto para describir la fuente de energía para la síntesis química de ATP. En términos más generales es utilizado para caracterizar la tendencia de difusión de los solutos a través de una membrana, un proceso que no implica una transformación química.
Gradientes iónicos[editar]
Con respecto a una célula, un orgánulo o un compartimento subcelular, la tendencia de un soluto con carga eléctrica, como un ion potasio K+, para moverse a través de la membrana, se decide en función de la diferencia de potencial electroquímico a ambos lados de la membrana, teniendo en cuenta tres factores:
- La diferencia en la concentración de soluto a ambos lados de la membrana.
- La valencia de la molécula de soluto.
- La diferencia de voltaje entre ambos lados de la membrana (potencial de membrana o Vm).
La diferencia de potencial electroquímico de un soluto es igual a cero en su “potencial de inversión”, voltaje transmembrana en el que el flujo neto del soluto a través de la membrana es también igual a cero. Este potencial se obtiene teóricamente de dos maneras posibles: o bien por la ecuación de Nernst (para los sistemas de una especie de iones permeables) o por la ecuación de Goldman-Hodgkin-Katz (utilizado cuando hay presencia de más de una especie de iones permeables). El potencial electroquímico se mide en el laboratorio utilizando electrodos de referencia.
A menudo se utilizan ATPasas transmembrana o proteínas transmembrana con dominios ATPasa para la elaboración y la utilización de gradientes iónicos. La enzima Na+/K+ ATPasa utiliza ATP para crear un gradiente de iones de sodio y un gradiente de iones de potasio. El potencial electroquímico es utilizado para almacenar energía. El acoplamiento quimiosmótico es una de las múltiples maneras por las que una reacción termodinámicamente desfavorable puede ser conducida por una reacción termodinámicamente favorable. El cotransporte de iones por simportadores y antiportadores es comúnmente utilizado para mover de forma activa iones a través de las membranas.
Gradientes de protones[editar]
El gradiente de protones puede ser utilizado como medio de almacenamiento energético para la producción de calor y rotación flagelar. Además, es una forma de energía interconvertible en transporte activo, generación de potencial electrónico, síntesis de NADPH y síntesis/hidrólisis de ATP.
La diferencia de potencial electroquímico a ambos lados de la membrana en mitocondrias, cloroplastos, bacterias y otros compartimentos de membrana que realizan transporte activo utilizando bombas de protones, es a veces llamada potencial quimiosmótico o fuerza motriz de protones (ver quimiosmosis). En este contexto, los protones se suelen considerar por separado utilizando las unidades de concentración o de pH.
Fuerza motriz protónica
En cada lugar de acoplamiento son expulsados dos protones, generando la fuerza motriz de protones (PMF). El ATP se crea indirectamente utilizando el PMF como fuente de energía. Cada par de protones produce una molécula de ATP.
Algunas arqueas (las más notables entre ellas son las halobacterias), crean gradientes de protones mediante el bombeo de protones del medio ambiente con la ayuda de la enzima bacteriorodopsina solar impulsada, que se utiliza en este caso para impulsar el motor molecular enzima ATP sintetasa para hacer los cambios necesarios en la conformación para que pueda ser sintetizado ATP. Las bacterias pueden crear también gradientes de protones mediante la utilización de la ATLP sintetasa en sentido inverso, y se utilizan para conducir los flagelos.
El F1F0 ATP sintetasa es una enzima reversible. Cantidades de ATP suficiente hacen que cree un gradiente de protones a ambos lados de la membrana. Este proceso es utilizado por bacterias que realizan la fermentación, las cuales no tienen una cadena de transporte electrónica, e hidrolizan ATP para crear un gradiente de protones (para los flagelos y el transporte de nutrientes al interior de la célula).
En bacterias que respiran bajo condiciones fisiológicas, la ATP sintetasa suele ir en dirección opuesta, creando ATP al tiempo que utiliza la fuerza motriz protónica creada por la cadena de transporte de electrones como fuente de energía. El proceso global de creación de energía de esta manera se llama fosforilación oxidativa. El mismo proceso tiene lugar en las mitocondrias, donde la ATP sintetasa se encuentra en la membrana mitocondrial interna, de modo que la parte F1 se une a la matriz mitocondrial, donde tiene lugar la síntesis de ATP.
El modelo de FitzHugh-Nagumo (FHN) describe un prototipo de un sistema excitable (por ejemplo, una neurona). Toma su nombre de Richard FitzHugh (1922 - 2007), quien propuso el modelo teórico en 1961, así como de J. Nagumo y otros, que construyeron un circuito electrónico equivalente.
El modelo FHN es un ejemplo de oscilador excitable ya que si el estímulo externo sobrepasa cierto valor umbral, el sistema exhibirá una excursión característica en el espacio de fases, tras la cual las variables y volverán a su valor en reposo.



Este comportamiento se observa en neuronas cuando son estimuladas por una corriente externa.
Matemáticamente, el sistema se expresa así:


El modelo FitzHugh–Nagumo es una versión simplificada del modelo de Hodgkin y Huxley, que describe la dinámica de la neurona más en detalle. En los artículos originales de FitzHugh, su modelo recibía el nombre de oscilador de van der Pol (Véanse Karl Friedrich Bonhoeffery Balthasar van der Pol). Esto es así porque el modelo de FitzHugh–Nagumo, para el caso , se reduce al oscilador de van der Pol.

El circuito equivalente fue propuesto por Jin-ichi Nagumo, Suguru Arimoto, y Shuji Yoshizawa.
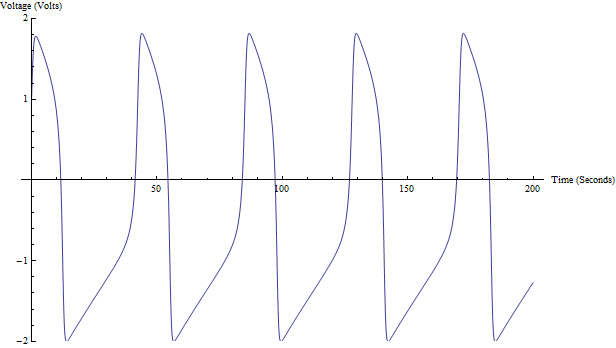
Representación de v con los parámetros I=0.5, a=0.7, b=0.8, y τ=12.5

En azul: trayectoria del modelo FHN en el espacio de fases. En rosa: nullcline cúbica. En amarillo: nullcline lineal.
El modelo de Hodgkin y Huxley describe cómo se inician y transmiten los potenciales de acción en las neuronas. Consiste en un conjunto de ecuaciones diferenciales ordinarias no lineales que aproxima las características eléctricas de células excitables como las neuronas o los miocitos cardíacos.
Alan Lloyd Hodgkin y Andrew Huxley propusieron el modelo en 1952 para explicar los mecanismos iónicos que subyacían a la iniciación y propagación de los potenciales de acción del axón gigante de calamar. En 1963recibieron el Premio Nóbel en fisiología - medicina por este trabajo.
Componentes básicos[editar]
- La membrana lipídica está representada como una capacitancia (Cm).
- Los canales iónicos están representados por una conductancia iónica no lineal (gn, donde n es un canal iónico específico), lo que significa que la conductancia depende del tiempo y del voltaje.
- Las puertas de los canales iónicos están representadas por conductancias lineales (gL).
- Los gradientes electroquímicos que conducen el flujo de iones están representados por baterías (En y EL), cuyos valores están determinados por el potencial de Nernst de la especie iónica de interés.
- Las bombas iónicas están representadas por fuentes de corriente (Ip).
La derivada en el tiempo del potencial a lo largo de la membrana () es proporcional a la suma de las corrientes en el circuito:


Donde Ii denota las corrientes iónicas individuales del modelo.
El modelo de FitzHugh-Nagumo es una simplificación del modelo de Hodgkin y Huxley.
No hay comentarios:
Publicar un comentario