crenación es el fenómeno en donde la célula animal se somete a una solución hipertónica. Al estar en esta solución con gran cantidad de soluto, tiende a liberar su agua. La destrucción de la célula se produce por deshidratación.
La crenación puede ser una característica de los glóbulos rojos de la sangre. Estos eritrocitos parecen tener proyecciones que se extienden desde una zona central más pequeña, como una bola de púas. Las crenaciones pueden ser o bien espículas de acantocitos grandes, irregulares, o proyecciones más pequeñas, irregulares de equinocitos. Acantocitos y equinocitos pueden surgir de anormalidades de los lípidos o proteínas de las membranas celulares, o de otros procesos de enfermedad, o como un artefacto ex vivo.
En la célula vegetal, el fenómeno es llamado plasmólisis y en este caso la deshidratación ocurre debajo de la pared celular.
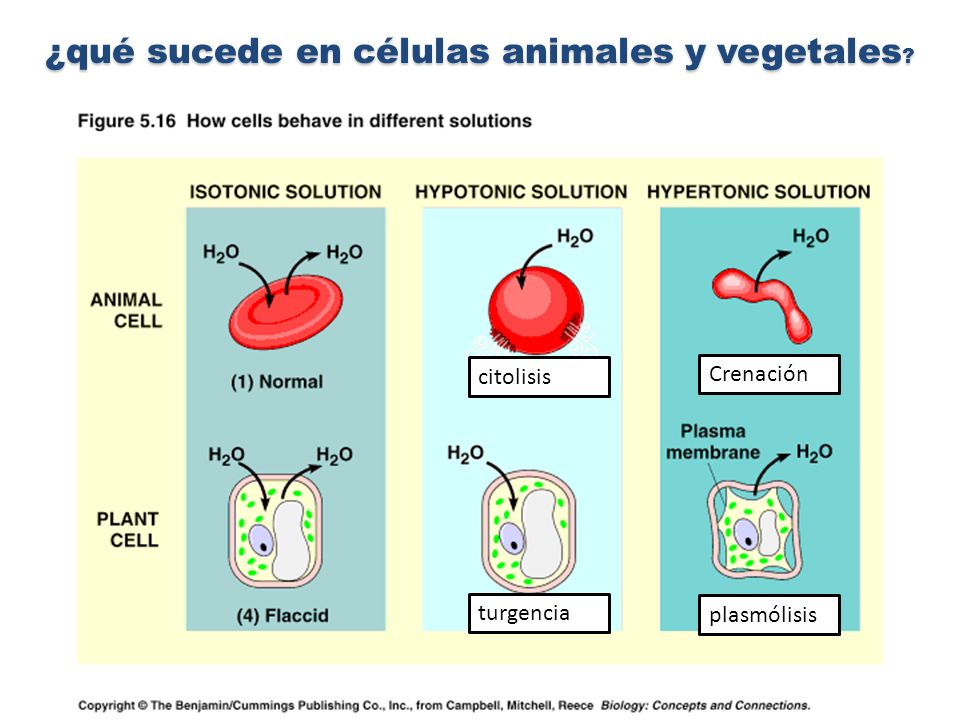
plasmólisis citolisis Crenación ...
diferenciación celular es el proceso por el cual las células de un linaje celular concreto (el linaje celular se determina en el momento de la formación del embrión) sufren modificaciones en su expresión génica, para adquirir la morfología y las funciones de un tipo celular específico y diferente al resto de tipos celulares del organismo.1
Cualquier célula que presente potencia (capacidad de diferenciación) es lo que se denomina célula madre. Estas pueden clasificarse según su capacidad de diferenciación en totipotentes, pluripotentes, multipotentes y unipotentes.
En los mamíferos, solo el cigoto y las células embrionarias jóvenes son totipotentes, mientras que en las plantas y hongos, muchas células son totipotentes.
Los últimos avances científicos están consiguiendo inducir a células animales diferenciadas para que pasen a ser totipotentes.
En los mamíferos, solo el cigoto y las células embrionarias jóvenes son totipotentes, mientras que en las plantas y hongos, muchas células son totipotentes.
Los últimos avances científicos están consiguiendo inducir a células animales diferenciadas para que pasen a ser totipotentes.

Diagrama de la división y diferenciación celular de la célula madre. A - célula madre; B - célula del progenitor; C - célula diferenciada; 1 - división simétrica de la célula madre, 2 - división asimétrica de la célula madre, 3 - división de la célula del progenitor; 4 - diferenciación terminal.
Introducción[editar]
En la inmensa mayoría de los organismos pluricelulares, todas las células no son idénticas. Por ejemplo, las células que forman la piel en el ser humano son diferentes de las células que componen los órganos internos. Sin embargo, todos los diferentes tipos celulares derivan de una sola célula inicial o cigoto, procedente de la fecundación de un óvulo por un espermatozoide, gracias a la diferenciación celular.
La diferenciación es un mecanismo mediante el cual una célula no especializada sufre modificaciones citológicas, dando lugar a los numerosos tipos celulares que forman el cuerpo como los miocitos (células musculares), los hepatocitos (células del hígado) los enterocitos (células del intestino) o incluso las neuronas(células del sistema nervioso).
La diferenciación es un mecanismo mediante el cual una célula no especializada sufre modificaciones citológicas, dando lugar a los numerosos tipos celulares que forman el cuerpo como los miocitos (células musculares), los hepatocitos (células del hígado) los enterocitos (células del intestino) o incluso las neuronas(células del sistema nervioso).
Durante la diferenciación, ciertos genes son expresados mientras que otros son reprimidos. Este proceso es intrínsecamente regulado gracias a distintos mecanismos de regulación de la expresión genética de las células. Así, la célula diferenciada expresará ciertos genes y adquirirá determinadas funciones.
La diferenciación metabólica, la sensibilidad a ciertas señales y la expresión de genes. Todos estos aspectos pueden ser modificados durante la diferenciación. En citopatología, el nivel de diferenciación celular es utilizado como una medida de la progresión de un cáncer.
Hipótesis sobre el mecanismo de diferenciación celular[editar]
Hasta la década de 1950, se planteaban dos posibles hipótesis que podrían explicar la diferenciación celular en los organismos pluricelulares. Una de ellas, es que a partir del embrión, los distintos tipos celulares perdían genes, regiones de su genoma, de forma que en el individuo adulto los distintos tipos celulares presentaran distinto genoma. La otra, defendía que manteniendo todos los tipos celulares el mismo genoma, existía una expresión diferencial de los distintos genes según el tipo celular.
A finales de los años 1950, Frederick Stewart cultivó células individuales de zanahoria en un medio con nutrientes y varias hormonas de crecimiento. El resultado es que algunas de ellas dieron lugar a zanahorias adultas completas. De esta forma se descartaba la hipótesis de la pérdida de material genético según el tipo celular
Mecanismos generales de control de la diferenciación celular[editar]
Como cualquier proceso celular, la diferenciación celular se debe a reacciones bioquímicas que tienen lugar en el interior de la célula, y está promovida por complejas cascadas de señalización.
Cabe destacar la importancia de las sustancias denominadas morfógenos. Estos son sustancias, normalmente proteínas que aparecen en un gradiente de concentración en la célula o en el medio que la rodea, de forma que controla el destino durante la diferenciación. Estos morfógenos serán clave en la señalización que lleve a la expresión de unos u otros genes.
La diferenciación celular, al igual que otros tantos procesos celulares, están controlados por mecanismos de regulación génica como control genómico, control transcripcional, control posttranscripcional, control traduccional y control posttraduccional.
Importancia de la impronta genómica[editar]
La impronta genómica es la expresión diferencial del alelo paterno o materno de un mismo gen debido simplemente a su procedencia. Este fenómeno juega un importante papel en el proceso de diferenciación celular.
Un ejemplo claro del importante papel que juega este fenómeno en la diferenciación celular se puede observar en el trabajo de J.A. Uranga (1994).
La partenogénesis es un fenómeno por el cuál algunos animales pueden reproducirse sin contribución de gametos masculinos. Sin embargo, a diferencia de en animales inferiores, en mamíferos la impronta genómica juega un papel muy importante y que se traduce en la inactivación de algunos genes.
Durante la implantación es donde se produce una gran mortandad de los embriones mamíferos activados partenogénicamente. J.A. Uranga utilizó un método de activación partenogénica que permite que la proporción de embriones partenogénicos que superan la implantación y llegan a la etapa de blastocito fuera similar al de embriones fecundados normalmente, y analizó las manifestaciones más tempranas de la impronta y su significado a nivel citológico y molecular.
Se observó que, aunque las exigencias metabólicas eran similares, en partenogénesis se inhibe la proliferación celular, de forma que el número de células es menor que en embriones fecundados.
Además se comprobó que existía una desregulación en la expresión de las citoqueratinas ya que se expresaban en las células indiferenciadas del interior del blastocito, cosa que no sucedía en los embriones fecundados. Este error en la expresión proteica se asocia a algún factor de origen paterno que inhibe la diferenciación de estas células.
Diferenciación celular en invertebrados[editar]
Ya que Drosophila melanogaster es un organismo modelo, su desarrollo embrionario con sus mecanismos de diferenciación están bien estudiados, y se describen correctamente en su artículo.

{kind=link}
Diferenciación celular en mamíferos[editar]
Los mecanismos de diferenciación celular en mamíferos se conocen menos, debido a los problemas que plantea la bioética en este campo. Sin embargo poco a poco nuestro conocimiento acerca de estos mecanismos es mayor. Un ejemplo puede ser el de los mioblastos C2C12 de ratones.
El mioblasto es el tipo celular precursor de los miocitos (células musculares), que dará lugar a éstas por diferenciación celular. Se ha observado en mioblastos C2C12 de ratón que la PLC-gamma 1 (phospholipase C-gamma) está relacionada con el proceso de diferenciación celular de estas células inducido con insulina. GC. Gaboardi et all (2010) para identificar las dianas corriente abajo de PLC-gamma 1 analizaron la expresión de isoformas de PKC (Phosphokinase C) dependientes de DAG (diacilglicerol) durante la diferenciación muscular. Se observó que durante la formación de miotubos, aumenta la expresión de PKC epsilon y PKC eta, y que PKC epsilon es capaz de formar un complejo con PLC-gamma 1. El aumento de la cantidad de PKC epsilon está asociado a un aumento de su actividad.
Analizaron la relación existente entre la cantidad de PKC epsilon y la expresión de miogenina. La conclusión fue, por tanto, que PKC epsilon desarrolla un papel muy importante en la diferenciación del músculo esquelético.
Diferenciación celular en plantas superiores[editar]
La diferenciación en plantas superiores se produce a partir de las células meristemáticas que son reclutadas para dar lugar a las células maduras que forman parte de los órganos de la planta. Los cambios que se producen en la célula afectan desde al contenido celular o estructura de la pared, hasta a las relaciones entre células vecinas (espacios entre células o crecimiento diferencial de unas respecto a otras).
Está demostrado que los genes de la familia WOX están relacionados con la organización de grupos de células durante el desarrollo de la planta. Según estudios realizados en el desarrollo de Arabidopsis thaliana y Solanum lycopersicum en los que se observó la transcripción y función de los genes WOX4, se constató que estos genes están involucrados en el desarrollo de los haces vasculares de la raíz y en el brote de los órganos laterales en ambas especies. Una reducción de la expresión de WOX4 mediante RNA de interferencia en Arabidopsis tuvo como consecuencia plantas de pequeño tamaño, cuyo floema y xilema no se había diferenciado o lo habían hecho dando lugar a conductos más pequeños de lo normal. Los datos obtenidos, sugieren que los genes WOX4 promueven la diferenciación o la no diferenciación del procambium vascular.
Reprogramación de células animales diferenciadas a células indiferenciadas[editar]
La clonación de la oveja Dolly demostró que en los ovocitos de mamífero se encontraban presentes ciertos factores de transcripción capaces de reprogramar el núcleo, no sólo manteniendo su estado de indiferenciación, sino induciendo en núcleos de células diferenciadas una vuelta hacia el estado indiferenciado.
Mediante estudios por técnicas de microarrays se encontraron algunos de estos factores como Oct4, Sox 2, Nanog, Tdgf1, Utf1, Lin28, etc. El funcionamiento de algunos de ellos ya se conoce bastante bien.
El hecho de que estos factores pueden no sólo mantener la célula indiferenciada si no reprogramar su núcleo una vez diferenciada, fue demostrado por el grupo de Yamanaka que fue capaz de encontrar una combinación de cuatro de estos factores capaz de reprogramar fibroblastos murinos: Oct2, Sox2, c-Myc y Klf4. También se ha conseguido reprogramar fibroblastos humanos con estos factores.
Debido a que c-Myc es un factor que es oncogénico, se ha seguido trabajando y se han encontrado nuevas combinaciones de cuatro factores en las que no aparecía c-Myc, lo cual nos puede mostrar la potencialidad que tiene esta línea de investigación, y la gran cantidad de rutas bioquímicas implicadas en el desarrollo celular.
DIFERENCIACIÓN CELULAR
La transformación morfológica y fisiológica de las células meristemáticas en tejidos adultos o diferenciados constituye el proceso de diferenciación celular. Ésta, y la consecuente especialización de la célula traen consigo la división de trabajo, formando células con funciones específicas. La diferenciación se produce por la activación diferencial de algunos genes y la represión de otros.
|
Durante el proceso de diferenciación las células sufren una serie de cambios en sus características y se produce un reajuste en sus relaciones mutuas. Los cambios principales son:
1) Alteraciones en el contenido celular (vacuolas, plástidos, sustancias ergásticas, alteraciones profundas del protoplasma o desaparición del mismo).
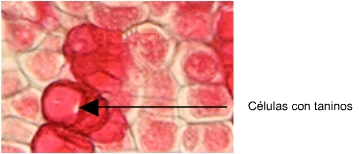 |
Imagen extraida de http://www.sbs.utexas.edu/mauseth/weblab
|
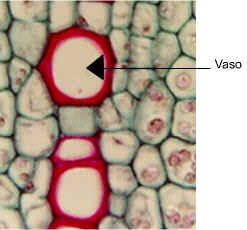 |
Imagen de http://www.sbs.utexas.edu/mauseth/weblab
|
Los espacios intercelulares pueden tener formación esquizógena o lisígena. En el primer caso se disuelve la laminilla media en los ángulos y aristas de las células. Como consecuencia, las células contiguas se separan en dichos lugares formando los espacios intercelulares o meatos, que pueden ensancharse por división de las células contiguas y formar cámaras mayores.
Cuando hay formación lisígena los huecos en los tejidos se originan por rotura de las células o disolución de las paredes celulares.
Espacios esquizógenos
| Espacios lisígenos |
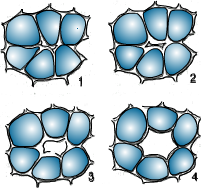 |  |
Imágenes modificadas de Fahn
| |
CRECIMIENTO SIMPLÁSTICO E INTRUSIVO.
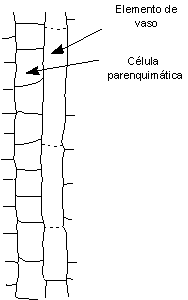
Se habla de crecimiento simplástico cuando el crecimiento de una célula se produce al unísono con el de las células vecinas. En cambio, hay crecimiento intrusivo cuando es desparejo con las células vecinas, cuando el elemento se abre camino entre ellas; las paredes de las células contiguas se separan como durante la formación de espacios intercelulares. Los plasmodesmos que existían desaparecen y generalmente no se establecen nuevas conexiones intercelulares en las porciones celulares que crecen intrusivamente. Los elementos alargados como las fibras y las traqueidas crecen por crecimiento intrusivo.
Crecimiento simplástico en corte longitudinal de raíz de Typha (totora)
| |
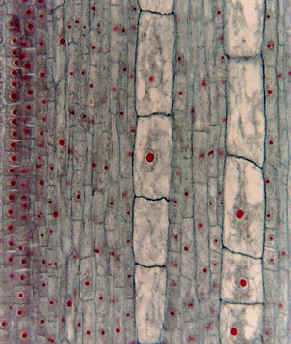 |  |
| Imagen extraida de http://www.sbs.utexas.edu/mauseth/weblab |
Imagen tomada de Esau 1974
|
| Crecimiento intrusivo de fibras a partir de células del cámbium |
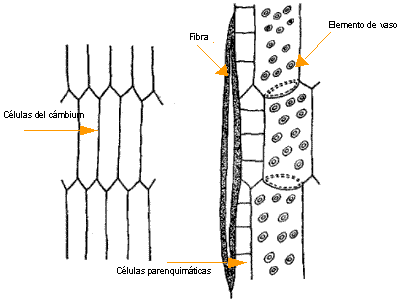 |
Imagen tomada de Esau 1974
|
No hay comentarios:
Publicar un comentario