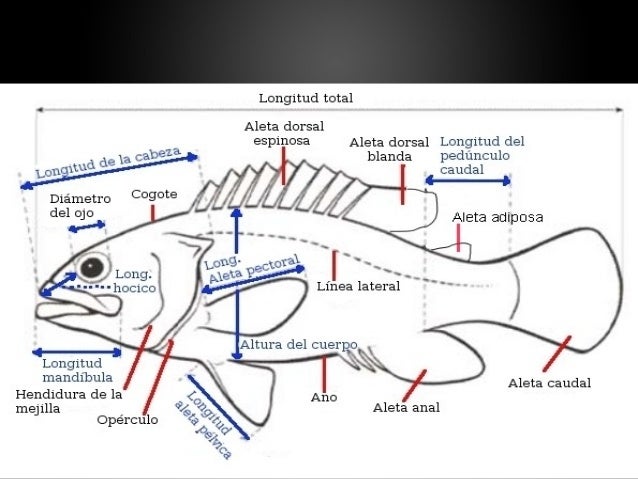
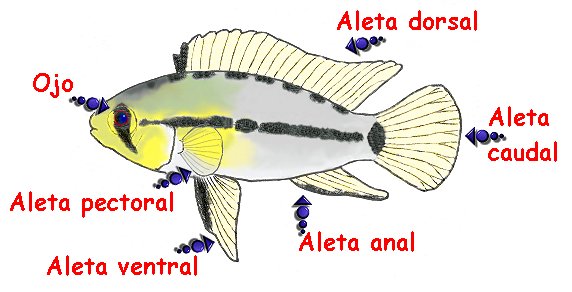
Las aletas son los órganos externos que muchos animales acuáticos utilizan para la locomoción y el equilibrio.
Tipos de aletas[editar]
2.png)
Pares[editar]
Son aquellas aletas que se encuentran fuera del plano de simetría bilateral, por lo que existe una en cada uno de los lados.
Pectorales[editar]
Se ubican detrás de las aberturas de las branquias. y ayudan al movimiento
Pélvicas o ventrales[editar]
Inmediatamente en frente de la parte posterior.
Esas aletas tienen una relación evolutiva con las extremidades de los vertebrados terrestres. Su función es principalmente estabilizadora y la de formar un disco succionador que puede ser usado para adherirse a objetos.
Impares[editar]
Son las que se ubican en el plano de simetría bilateral. El número de ellas no es necesariamente impar (puede haber, por ejemplo, dos aletas dorsales).
Dorsales[editar]

Una aleta dorsal es una aleta localizada en la espalda de algunos peces, ballenas, delfines, marsopas.
Su propósito principal es estabilizar al animal contra el rollido y asistir en súbitos giros. Algunos animales han desarrollado aletas dorsales con funciones protectoras, como las espinas o venenos. Muchos siluros pueden trabar su espina central de la aleta dorsal en una posición extendida para asegurar su predación o clavarlos contra una grieta.
Las espinas dorsales se presentan en una variedad de formas y tamaños. Y son famosas en mostrarse como signo de acercamiento de tiburones.
Ballenas, delfines, y marsopas[editar]
En cuanto al tamaño, una aleta dorsal de una orca macho es de 1,8 metros de alto. (La orca hembra lo tiene más corto y más curvada.) Muchas (30-100%) orcas machos en cautividad experimentan el colapso de la aleta dorsal, posiblemente por falta de ejercicios y decayendo el tono muscular, otros posibles factores pueden ser cambio de dieta, navegación zigzageante, reacción con el nitrato de sodio en la glándula pituitaria perdiendo entonces tejido conectivo de la aleta dorsal (pero solo cerca del 1% de las orcas salvajes colapsan sus aletas dorsales.) Las aletas dorsales de muchas otras ballenas son relativamente pequeñas. La ballena boreal Balaena mysticetus no tiene aleta dorsal, como adaptación evolucionaria de su vida navegando debajo de los hielos.
Las aletas dorsales de las ballenas, con el tiempo, desarrollan marcas y patrones distintivos, ayudando a los biólogos a identificar individuos en el campo.
Caudal[editar]

A veces lobulada, en la parte posterior del cuerpo. Tiene radios espinosos y simples pero también pueden ser compuestos. Se clasifican en:
- Heterocerca
- Protocerca
- Homocerca
- Dificerca
Anal[editar]
Se ubican en la región ventral, detrás del ano.
Estas aletas son excrecencias de la piel y en los peces teleósteosson soportadas por radios, que son escamas modificadas. En los peces cartilaginosos (Chondrichthyes), son soportadas por filamentos de ese material.
En los mamíferos acuáticos, como las ballenas y las focas, lo mismo que en las aves (pingüinos) las aletas son transformaciones de las terminaciones de sus extremidades (manos y pies).

Aleta adiposa[editar]
La aleta adiposa es una aleta de consistencia carnosa encontrada entre la aleta dorsal y la aleta caudal. Se encuentra en nueve órdenes de peces teleósteos, en percopsiformes, myctophiformes, aulopiformes, stomiiformes, salmoniformes, osmeriformes, characiformes, siluriformes y argentiniformes. Su tamaño es muy variable.
Fórmula de las aletas[editar]
Es usada por los ictiólogos para describir un pez y reconocer las diferencias entre especies. En general, la aleta es identificada mediante letras mayúsculas (D: dorsal; A: anal; T: torácica; AB: abdominal; C: caudal), el número de radios espinosos se escribe en cifras romanas, y el número de radios blandos en cifras arábigas.
Por ejemplo:
- D IV/8, A III/5 = Aleta dorsal con 4 espinas y 8 radios blandos, aleta anal con 3 espinas y 5 radios blandos.
- D2 3/9 = Segunda dorsal con 3 radios blandos indivisos y 9 radios blandos divididos.

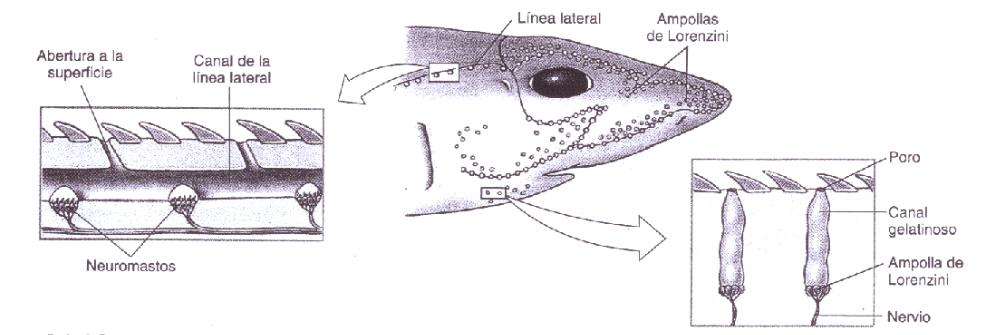
Las ampollas de Lorenzini son órganos sensorialesespeciales, formados por una red de canales con electrorreceptores cubiertos con una sustancia gelatinosa, encontrados en los Condrictios (tiburones, rayas y quimeras). El italiano Marcello Malpighi descubrió las ampollas de Lorenzini, que serían descritas por primera vez de modo detallado en 1678 por el médico italiano Stefano Lorenzini. La importancia de estas radica en detectar los campos electromagnéticos. Los tiburones, por ejemplo, las usan para encontrar a sus presas, para seguir la corriente del agua puesto que si van en contra de ella se cansaran fácilmente.
Características generales[editar]
Cada ampolla consiste en un canal lleno de sustancia gelatinosa en cuyo fondo se encuentran grupos de pequeñas bolsas llenas de células electrorreceptoras, con una abertura hacia el exterior por medio de un poro en la piel. Las ampollas generalmente están agrupadas en el cuerpo, cada grupo tiene ampollas conectadas con diferentes sitios de la piel, preservando la simetría bilateral. Los largos de los canales varían de un animal a otro, pero la distribución de los poros es aproximadamente específica para cada especie. Los poros de las ampollas son claramente visibles como puntos oscuros en la piel. Proveen a los elasmobranquios de un "sexto sentido" capaz de detectar tanto campos electromagnéticos como gradientes de temperatura.
Funciones[editar]
A comienzos del siglo 20, no estaba claro el propósito de las ampollas, y experimentos electrofisiológicos sugirieron una sensibilidad a la temperatura, presión mecánica y quizás salinidad. No fue hasta 1960 que las ampollas fueron claramente identificadas como órganos receptores especializados en sentir campos eléctricos. Las ampollas podrían también permitir detectar cambios en la temperatura del agua. Cada ampolla es un fajo de células sensoriales que contienen múltiples fibras nerviosas. Estas fibras encerradas en un túbulo de sustancia gelatinosa que tiene una abertura directa a la superficie a través del poro. El gel es una sustancia basada en glicoproteína con la misma resistencia que el agua marina, y tiene propiedades eléctricas similares a un semiconductor, permitiéndole esencialmente transducir los cambios de temperatura a una señal eléctrica que podría ser utilizada para detectar gradientes de temperatura.
Las ampollas detectan campos eléctricos en el agua, o más precisamente la diferencia entre el voltaje en el poro y el voltaje en la base del electrorreceptor. Un estímulo positivo del poro disminuiría la tasa de actividad del nervio proveniente de las células electrorreceptoras, y un estímulo negativo del poro aumentaría la tasa de actividad proveniente de las células electrorreceptoras.
Los tiburones podrían ser más sensitivos a los campos eléctricos que cualquier otro animal, con un umbral de sensibilidad tan bajo como 5nV/cm. Esto es 5/1,000,000,000 de un voltio medido en un centímetro de largo de ampolla. Ya que todos animales producen un campo eléctrico en la contracción muscular, es fácil imaginar que el tiburón podría captar estímulos eléctricos débiles emitidos particularmente por sus presas. Por otra parte, en tanques experimentales, los campos electroquímicos generados por presas paralizadas serían suficiente para producir el ataque por parte de tiburones y rayas, por tanto las contracciones musculares no son lo único necesario para atraerlos.
Los campos eléctricos producidos por las corrientes oceánicas que se mueven en el campo electromagnético de la Tierra son del mismo orden de magnitud que los campos eléctricos que los tiburones y rayas son capaces de sentir. De esta manera, se pueden orientar por los campos eléctricos de las corrientes oceánicas, y usar otras fuentes de campos eléctricos en el océano para su orientación local. Adicionalmente, el campo eléctrico que producen cuando nadan en el campo magnético de la Tierra podría darles pistas sobre su dirección magnética.

Electrorreceptores (Ampolla de Lorenzini) y los canales de la línea lateral en la cabeza de un tiburón.
aparato de Weber o aparato weberiano es una estructura anatómica que conecta la vejiga natatoria al sistema auditivo en los peces pertenecientes al Super Orden de los Ostariophysi. Cuando se encuentra completamente desarrollado en un pez adulto, los elementos del aparato son considerados en conjunto como los huesesillos de Weber. La presencia de ésta estructura es una de las características más importantes y filogenética que distingue al grupo Ostariophysi. La estructura consiste en un conjunto de huesos diminutos que se originan de las primeras vértebras para desarrollar un embrión de esta especie. Estos huesos crecen para conectar físicamente el sistema auditivo, específicamente el oído interno a la vejiga natatoria.1 2 La estructura actúa como un amplificador de ondas de sonido, de otro modo sería sólo ligeramente perceptible por la estructura del oído interno.
Anatomía estructural y funcionamiento[editar]
La estructura generalizada del aparato de Weber es parecida a un complejo esquelético de huesos y huesecillosque están conectados físicamente al complejo laberinto auditivo anterior y a la región de la vejiga natatoria posterior. La estructura completa es derivada de elementos del esqueleto de las primeras cuatro vértebras. Los elementos involucrados son: los huesos supraneurales del cráneo; huesos neuronales en forma de arcos modificados; el claustrum, tripus, intercalarium y el scaphium (huesecillos de Weber); el os suspensorium de la cuarta vértebra; la parapófisis de la quinta vértebra incluyéndola, más la costilla pleural. Además, una estructura compuesta por espinas neurales fusionadas forman la parte dorsal del aparato de Weber. En conjunto, la estructura interactúa con el otolito lagenar anterior dentro del cráneo y la vejiga natatoria posterior por medio de la costilla pleural. Postero-ventralmente, se encuentra el tripus, el os suspensorium y la tercera costilla que interactúa directamente con la cámara anterior de la vejiga natatoria.1
El aparato weberiano funciona transmitiendo señales auditivas desde la vejiga, a través de los huesecillos de Weber y después directo a la estructura del oído interno. Esencialmente, la estructura actúa como un amplificador de ondas de sonido con la función que añade la vejiga natatoria como una cámara de resonanciadonde amplifica la señal para ser audible.2
Embriología[editar]
El análisis embrionario del aparato de Weber de la taxonomía Brycon ha arrojado algo de luz al desarrollo de la estructura misma. Los elementos del aparato weberiano se forman de las primeras cinco vértebras distinguibles del individuo. Los supraneurales comienzan como un elemento del cráneo. La claustra y la scaphia se desarrollan a partir de elements expandidos del arco neural de la primera vértebra (V1). A partir de la segunda vértebra (V2), el intercalarum y el proceso lateral de la vértebra son reducidos y agrupados. La costilla pleural (R1) de la tercera vértebra (V3) se contrae y mueve un poco ventralmente, formando el tripus de un parapófisis vertebral fusionándose con la costilla pleural. El hueso os suspensorium de la cuarta vértebra (V4) permanece en su forma, desarrollándose de la costilla pleural de la vértebra (R2). Los elementos restantes de la quinta vértebra (V5), la parapófisis y la costilla de articulación (R3), incluyendo la vértebra misma, forman la estructura posterior del aparato de Weber. Las espinas neurales de las primeras cuatro vértebras se fusionan y comprimen, formando una de las mayores estructuras del aparato.1 2
Es estudio de la embriología del aparato de Weber ha sido llevado a cabo en varias otras especies del orden Ostariophysi. Los resultados han dado lugar a diversas interpretaciones del desarrollo (por lo tanto la homología) de las estructuras que forman al aparato. Estudios específicos han sido realizados sobre el aparato weberiano de unos pocos taxones seleccionados, incluyendo Danio rerio,3 Rhaphiodon vulpinus4 y Corydoras paleatus.5
Historia de su evolución[editar]
La incidencia más temprana registrada de un aparato weberiano es de fósiles de peces de los Santanichthys diasii, datos de principios del Cretáceo al noreste de Brasil. En el taxón citado, el aparato de Weber está bastante desarrollado; teniendo un intercalarium distinguible y un tripus articuladp a una segund ay tercera vértebra respectivamente. Inlcuos una scaphia puede observarse en al menos dos especímenes. El arco neura de la tercera vértebra, se encuentra ampliado, casi similar a los especímenes modernos. El claustrum, elemento del aparato de Weber moderno, no se encuentra en la especie Santanichthys diasii. Solo las primeras cuatro vértebras están involucradas en el aparato de Weber del Santanichthys diasii, y no hay rastros de la participación de la quinta vértebra como en los especímenes modernos.6
Etimología[editar]

2.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
El aparato de Weber lleva el nombre del anatomista y fisiólogo alemán Ernst Heinrich Weber (1795 – 1878). El aparato fue descrito por primera vez en detalle por Weber. Solo cuatro huesos fueron identificados, específicamente el claustrum, scaphium, intercalarium y el tripus. En conjunto, estos elementos fueron hipotetizados en desempeñar un papel en las funciones auditivas.7 A través de los años, se han propuesto y descartado otras funciones, como la regulación hidrostática, que fue una de las primeras sugerencias para la función del aparato.
No hay comentarios:
Publicar un comentario