difusión simple al proceso por el cual se produce un flujo de moléculas a través de una membrana permeable sin que exista un aporte externo de energía. Este proceso, que en última instancia se encuentra determinado por una diferencia de concentración molecular entre los dos medios separados por la membrana, no requiere de un aporte de energía debido a que su principal fuerza impulsora es el aumento de la entropía total del sistema.
En este proceso el desplazamiento de las moléculas se produce siguiendo el gradiente de concentración, las moléculas atraviesan la membrana desde el medio donde se encuentran en mayor concentración, hacia el medio donde se encuentran en menor concentración.
El proceso de difusión simple se encuentra descrito por las Leyes de Fick, las cuales relacionan la densidad del flujo de las moléculas con la diferencia de concentración entre los dos medios separados por la membrana, el coeficiente de difusión de las mismas y la permeabilidad de la membrana.
El proceso de difusión simple es de vital importancia para el transporte de moléculas pequeñas a través de las membranas celulares. Es el único mecanismo por el cual el oxígeno ingresa a las células que lo utilizan como aceptor final de electrones en la cadena respiratoria y uno de los principales mecanismos de regulación osmóticaen las células.

Generalidades[editar]
Difusión[editar]
Difusión es el proceso irreversible por el cual un grupo de partículas se distribuye de manera uniforme en un medio ya sea vacío o formado por otro grupo de partículas. Este proceso es estadísticamente predecible en conjunto, aunque el movimiento de cada partícula aislada es totalmente aleatorio. Se encuentra impulsado por el movimiento térmico de las partículas que componen ese sistema y se produce siguiendo las líneas de mayor diferencia de concentración entre regiones, esto es, siguiendo los gradientes de concentración.
En las membranas celulares[editar]
En el caso de las células vivas, el proceso de difusión simple se establece a través de la membrana celular, por lo que de hecho existen tres procesos de difusión encadenados, una difusión que ocurre en el medio de mayor concentración, una difusión que ocurre en el medio de separación y una difusión que ocurre en el medio de menor concentración. Como el proceso limitante de la velocidad es la difusión a través del medio de separación, se puede simplificar un modelo donde el flujo de partículas depende de la diferencia de concentración entre ambos lados del medio de separación y del tipo de interacciones que presente la molécula que va a atravesar la membrana con ese medio.
Las moléculas que pueden atravesar con facilidad las membranas celulares, debido a este fenómeno, son únicamente las de los gases (por ejemplo CO2, O2), las moléculas hidrofóbicas (por ejemplo benceno) y las moléculas polares pequeñas (por ejemplo H2O y etanol), esto es así debido a que las moléculas hidrofóbicas y apolares son solubles en la región central apolar de la bicapa lipídica, y las moléculas polares pequeñas son lo suficientemente pequeñas como para que las interacciones desfavorables se vean compensadas por un aumento de la entropía del sistema. Por otra parte, las moléculas polares grandes tales como la glucosa, los aminoácidos y las moléculas cargadas o iones (Hx+, Na+, Cl+ y Ca2+) establecen interacciones demasiado fuertes con el medio acuoso fuera de la bicapa lipídica, por lo que les resulta muy desfavorable desde el punto de vista energético romper estas interacciones para atravesar la región central hidrofóbica. Como consecuencia las membranas biológicas son prácticamente impermeables a este tipo de moléculas, por lo que requieren de otros mecanismos de transporte.r
Leyes de Fick en la difusión simple[editar]
Para el estudio del transporte a través de las membranas celulares por difusión simple, es necesario considerar las leyes que rigen los procesos de difusión: las Leyes de Fick. Cuando un sistema presenta una diferencia en el número de moléculas por unidad de volumen (concentración), por dentro y por fuera de un espacio delimitado por una membrana, se establece un gradiente de concentración que, expresado en forma diferencial sencilla, es proporcional a la diferencia en la concentración entre ambos medios (c) e inversamente proporcional al espesor de la membrana (x):

En caso de que la membrana sea permeable a las moléculas desigualmente distribuidas, se establece un flujo neto de partículas desde la zona de mayor concentración hacia la zona de menor concentración. La densidad de partículas en este flujo (J) depende del gradiente de concentración y de la facilidad con que las partículas atraviesan la membrana (D o coeficiente de difusión). El signo negativo indica la dirección del flujo (de mayor a menor concentración
(1) Primera Ley de Fick.

Al considerar una membrana de espesor no infinitesimal relativamente constante, en la cual se presenta una diferencia de concentración también constante, la primera Ley de Fick se puede reescribir como:


En esta ecuación el coeficiente de permeabilidad de la membrana queda definido como:

Por lo que la primera ley de Fick también puede escribirse cómo:

La ecuación de continuidad que expresa la conservación del número de moléculas, obtenida a partir del análisis del flujo entrante y saliente de las moléculas a través de un área y de la rapidez de acumulación (aumento por unidad de tiempo del número de partículas unidad de volumen) es:

Reemplazando, se obtiene:

Segunda ley de Fick (ecuación de difusión)[editar]
Se puede acordar un modelo matemático ideal que permite comprender el comportamiento de las moléculas y de las variables que intervienen en el proceso de difusión, pero la creación de este modelo requiere una serie de asunciones también ideales:
- Se asume un modelo de membrana de espesor constante, a través de la cual sólo se pueden difundir moléculas no polares pequeñas.
- Es necesario asumir que el gradiente de concentración varía de manera constante dentro de la membrana. A partir de ello y de la primera Ley de Fick, se halla la densidad de corriente de partículas J en función del gradiente de concentración que permite hacer un análisis de la dependencia mutua de estas dos variables.

- Para obtener la solución a la segunda Ley de Fick, es necesario asumir que en todo el sistema considerado y que en a tiempo cero (t = 0) se introduce instantáneamente en un punto del plano que corresponde a x = 0 una cantidad de sustancia igual a N moléculas/. Esto implica que cuanto más cercano es el tiempo a cero, la concentración también tiende a cero en todas partes menos en el punto de aplicación x=0.


Bajo estas condiciones la segunda Ley de Fick también debe satisfacer la condición de borde para y siendo la expresión final, obtenida a través de las transformadas de Fourier, la siguiente:




Ésta es la solución desarrollada de la segunda ley de Fick para una sola dimensión.
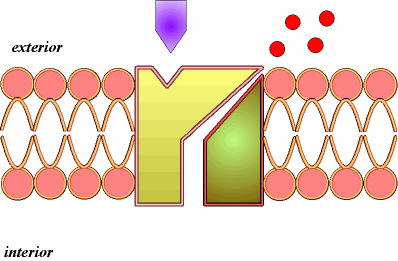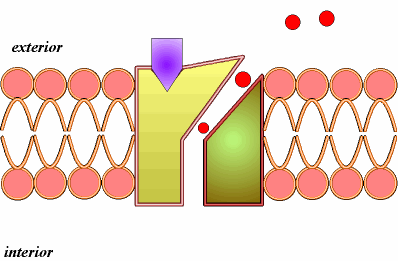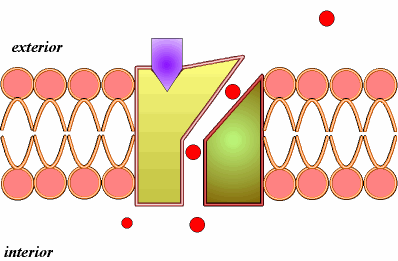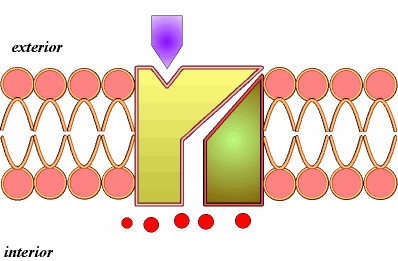
- Difusión simple . Es el paso de pequeñas moléculas a favor del gradiente; puede realizarse a través de la bicapa lipídica o a través de canales proteícos.
A) Difusión simple a través de la bicapa (1). Así entran moléculas lipídicas como las hormonas esteroideas, anestésicos como el éter y fármacos liposolubles. Y sustancias apolares como el oxígeno, el CO2 y el nitrógeno atmosférico. Algunas moléculas polares de muy pequeño tamaño, como el agua, el etanol y la glicerina, también atraviesan la membrana por difusión simple. La difusión del agua recibe el nombre de ósmosis
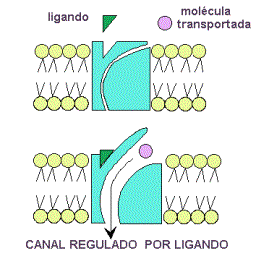
B) Difusión simple a través de canales (2).Se realiza mediante las denominadas proteínas de canal. Así entran iones como el Na+, K+, Ca2+, Cl-. Las proteínas de canal son proteínas con un orificio o canal interno, cuya apertura está regulada, por ejemplo por ligando, como ocurre con neurotransmisores u hormonas, que se unen a una determinada región, el receptor de la proteína de canal, que sufre una transformación estructural que induce la apertura del canal.
 | 
La endocitosis ( del griego ἐνδο- gr. 'dentro', kyto- κύτος gr. cient. 'célula' y -ō-sis gr. 'proceso') es un mecanismo clave por el cual las células introducen moléculas grandes, partículas extracelulares e inclusive pequeñas células, englobándolas en una invaginación de la membrana citoplasmática, formando una vesícula que termina por desprenderse de la membrana para incorporarse al citosol.Se presenta como un caso contrario a los acontecimientos de la exocitosis.12345
Los mecanismos de endocitosis tienen como fin regular la composición de lípidos y proteínas de la membrana plasmática, realizan una regulación de cómo las células interactúan con su entorno y por ende es un apoyo fundamental para la fisiología celular y la homeostasis.34
El término de "endocitosis" fue utilizado por Christian deDuve en 1963 como concepto para incluir en un mismo término a la ingestión de partículas grandes y la absorción de fluidos o macromoléculas en pequeñas vesículas.
Tipos de Endocitosis[editar]
Existen tres tipos de endocitosis:
Cuando la endocitosis da lugar a la captura de partículas se denomina fagocitosis ("la célula come"), y cuando son solamente porciones de líquido las capturadas, se denomina pinocitosis ("la célula bebe").2 La pinocitosis atrapa sustancias de forma indiscriminada, mientras que la endocitosis mediada por receptores sólo incluye al receptor y a aquellas moléculas que se unen a dicho receptor, es decir, es un tipo de endocitosis muy selectivo. En los tres casos la membrana plasmática se pliega hacia adentro formando una especie de bolsillo que dará lugar a una vesícula para la posterior migración de su contenido al interior de la célula.1
Fagocitosis[editar]
El mecanismo de endocitosis más común es la fagocitosis, este consiste en la introducción de un sólido al medio intracelular. Estos pueden ser: una molécula de gran tamaño, una partícula o un microorganismo, como por ejemplo; bacterias, polvo atmosférico, células o restos celulares.7
Primero la partícula se apoya en una zona de la membrana celular produciéndose una invaginación, al ingresar a la célula, que se estrangula, quedando lo ingresado envuelto en la membrana plasmática, constituyendo una vesícula denominada fagosoma, ésta es creada por medio del citoesqueleto. El fagosoma se fusiona con los lisosomas, formando un fagolisosoma, los orgánulos encargados de realizar la digestión celular. Generalmente es una función de defensa cuando se presentan agresiones del exterior o de renovación de tejidos orgánicos.78
Es importante saber que la unión de la partícula a los receptores en la superficie de la célula fagocítica desencadena la extensión de los pseudópodos, un movimiento basado en actina de la superficie celular; los pseudópodos rodean la partícula y sus membranas se fusionan para formar una gran vesícula intracelular (> 0,25 μm de diámetro) llamado fagosoma, estos se fusionan con los lisosomas, generando a los fagolisosomas, en los mismos donde el material ingerido es digerido por la acción de las hidrolasas ácido lisosómicas; durante la maduración del fagolisosoma, algunas de las proteínas de membrana internalizadas se reciclan a la membrana plasmática.6
Muchas amebas usan fagocitosis para capturar partículas de alimentos, como bacterias u otros protozoos; mientras tanto en animales multicelulares, las principales funciones de la fagocitosis consisten en proporcionar una defensa contra microorganismos invasores y eliminar células envejecidas o dañadas del cuerpo, específicamente en los mamíferos, la fagocitosis es la función de dos tipos de glóbulos blancos, macrófagos y neutrófilos, a los que frecuentemente se los llama como "fagocitos profesionales", poseen papeles sumamente importantes para los sistemas de defensa del cuerpo al eliminar los microorganismos de los tejidos infectados, además, los macrófagos, eliminan las células viejas o muertas de los tejidos en todo el cuerpo, son responsables de la eliminación de más de 1011 células sanguíneas envejecidas sobre una base diaria.6
Endosomas
Se encuentran dentro de la célula, son compartimientos intermedios, tienen la función de destruir materiales que han sufrido endocitosis, fagocitosis o autofagocitosis y para la formación de los lisosomas.9
Pinocitosis[editar]
Las células que llevan a cabo la pinocitosis presentan una región en la membrana plasmática que está recubierta por una proteína (la clatrina) en su cara citosólica, de forma que cuando la molécula se deposita sobre esa región de membrana se forma un caparazón revestido que la rodea, posteriormente perderá ese revestimiento para poder ser digerida por los lisosomas. En este proceso la célula incorpora líquidos en los cuales pueden estar disueltas proteínas.7
La parte endocítica del ciclo usualmente empieza en fosas recubiertas de clatrina, las mismas que generalmente ocupan alrededor del 2% del área total de la membrana plasmática, el tiempo de vida de estas fosas es corto (alrededor de un minuto después de ser formado), se invagina dentro de la célula y se aprieta para formar una vesícula revestida con clatrina; se estima que unas 2500 vesículas revestidas con clatrina abandonan la membrana plasmática de un fibroblasto cultivado cada minuto, estas vesículas son aún más transitorias que las cavidades revestidas, cuyo tiempo de vida es de unos los pocos segundos después de su formación, por lo que desprenden su capa y son capaces de fundirse con los endosomas tempranos; debido a que el fluido extracelular está atrapado en fosas recubiertas de clatrina, a medida que se invaginan para formar vesículas recubiertas, cualquier sustancia disuelta en el fluido extracelular se internaliza, proceso denominado endocitosis en fase fluida.10
La velocidad a la que la membrana plasmática realiza este proceso varía entre los tipos de células, pero suele ser increíblemente grande; por ejemplo un macrófago, ingiere el 25% de su propio volumen de líquido cada hora.10
Es importante conocer que existen otros mecanismos menos conocidos por los cuales las células pueden formar vesículas pinocíticas sin formar cavidades y vesículas revestidas con clatrina; una de estas vías se inicia en caveolae (vocablo latín que significa "pequeñas cavidades"), la misma que es reconocida por su capacidad para transportar moléculas a través de las células endoteliales que forman el revestimiento interno de los vasos sanguíneos, los caveolae están presentes en la membrana plasmática de la mayoría de los tipos de células, y en algunos de ellos se ven como frascos profundamente invaginados en el microscopio electrónico, se cree que se forman a partir de balsas lipídicas, parches de la membrana plasmática que son especialmente ricos en colesterol, glicoesfingolípidos y proteínas de membrana ancladas en GPI, su principal proteína estructural es la caveolina, proteína de membrana integral multipaso que es miembro de una familia heterogénea de proteínas.10
Se cree que las caveolae invaginan y recogen proteínas de carga en virtud de la composición lipídica de la membrana calveolar, en lugar del ensamblaje de un revestimiento proteico citosólico, se separan de la membrana plasmática y pueden entregar su contenido a compartimentos parecidos a endosomas o (en un proceso llamado transcitocis) a la membrana plasmática en el lado opuesto de una célula polarizada, algunos virus que atacan a animales entran en las células en vesículas derivadas de caveolae,10 produciéndose un ciclo infeccioso.10
Endocitosis Mediada por receptores[editar]
La endocitosis mediada por receptores de carga, se produce cuando los receptores se acumulan en regiones bien definidas de la membrana celular, este mecanismo de transporte permite la entrada selectiva de moléculas en la célula.11
El proceso contrario a la endocitosis es la exocitosis. La endocitosis y exocitosis son procesos de entrada y salida de sustancias que permiten mantener la homeostasis de la célula, están regulados por la misma con el fin de mantener constante la membrana plasmática, ya que permiten su regeneración pues los fagosoma que contienen las moléculas fagocitadas se forman a partir de la membrana plasmática y cuando el proceso de digestión celular llevado a cabo por los lisosomas finaliza se lleva a cabo la excreción celular por exocitosis recuperándose la membrana utilizada para la formación del fagosoma.12
La vesícula formada se llama endosoma que fusionará con un lisosoma donde se produce la digestión intracelular del contenido de esta.Las células fagocíticas especializadas presentan receptores de membrana que cuando contactan con fragmentos celulares inducen la formación de pseudópodos que la recubren formando los fagosomas. En la especie humana, las principales células con capacidad para fagocitar (fagocitos) son los polimorfonucleares neutrófilos (PMN), que son de vida corta. También participan los fagocitos mononucleares (SFM) constituido por los monocitos circulantes y los macrófagos tisulares.13
Posteriormente los lisosomas se fusionan con la pared de los fagosomas vertiendo sus enzimas hidrolíticas que actúan a pH ácido(próximo a 5) y llevan a cabo la degradación de los fragmentos celulares. Aquella parte que no puede ser digerida se eliminará al exterior mediante exocitosis en el proceso conocido como defecación celular.
Ejemplos
Endocitosis mediada por clatrina[editar]
Se produce en todas las clases de células de mamíferos y cumple funciones importantes como la absorción de nutrientes y la comunicación intracelular. Este proceso es el principal mecanismo de internalización de macromoléculas y componentes de la membrana plasmática. Se considera como un mecanismo de accionamiento que implica más de cincuenta componentes diferentes de proteínas de ensamblaje, restringiéndose a una sola ubicación en la membrana plasmática. Localizadas en una vía temporal ordenada y jerárquica.3
La mayoría de los virus que presentan envoltura hacen uso de un mecanismo de endocitosis con el fin de entrar en una célula permisiva e iniciar la infección. La endocitosis por clatrina es un campo de investigación que ha desatado gran interés en los últimos tiempos.14
Endocitosis mediada por caveolina[editar]
Es un proceso regulado por complejos de señalización a través de la GTPAasa. Esta vía es utilizada por patógenos para escapar de la degradación por enzimas lisosomales. Las caveolas son invaginaciones de la membrana en forma de botella, que tienen un tamaño entre 50 y 100 nm, las cuales están revestidas por caveolina.
Este proceso es fundamental ante una respuesta inmune, la comunicación intercelular, la transducción de señales, la homeostasis tanto celular como la del organismo completo; a principios de 2006 se encontró que una proteína marcadora de base lipídica, llamada flotilina-1, participa en una nueva vía de endocitosis.15
Las neuronas utilizan el mecanismo de endocitosis para recuperar un neurotransmisor liberado en el espacio sináptico, para ser reutilizado. Sin este proceso, se produciría un fracaso en la transmisión del impulso nerviosoentre neuronas.
Un ejemplo de endocitosis mediada por receptores es cuando las células humanas incorporan el colesterol el cual es utilizado en la síntesis de membranas y también como precursor de otros esteroides.16
Este proceso tiene una fuerte relación con la erradicación de la infección por VIH-1. La caveolina-1 mediada por la absorción de VIH-1 es un mecanismo de restricción intrínseca presente en las células de Langerhans en los humanos que previene esta infección. El aprovechar esta vía de internalización, tiene el potencial de elaborar estrategias para una lucha contra la transmisión del esta enfermedad.17
Las pequeñas invaginaciones de la membrana plasmática, caveolae, revestidas con caveolina, contienen algunas proteínas receptoras y se usan para ciertos tipos de endocitosis mediada por receptores, pero esta última se produce de forma usual a través de cavidades y vesículas recubiertas de clatrina, proceso es similar al envasado de las enzimas lisosómicas por la manosa 6-fosfato (M6P) en el trans-Golgi, la mayoría de los receptores M6P están localizados en el trans-Golgi, pero algunos se encuentran en la superficie celular y las enzimas lisosómicas que se secretan se unen a estos receptores y se devuelven a las células mediante este tipo de endocitosis; en otras palabras se puede afirmar que las proteínas del receptor transmembrana que se internalizan desde la superficie celular durante la endocitosis se clasifican y reciclan de nuevo a la superficie celular, de manera muy similar al reciclaje de los receptores M6P a la membrana plasmática y trans-Golgi.18
Algunos receptores se agrupan sobre fosas recubiertas de clatrina incluso en ausencia de ligando, otros se difunden libremente en el plano de la membrana plasmática pero sufren un cambio conformacional al unirse al ligando, de modo que cuando el complejo receptor-ligando se difunde en un pozo revestido con clatrina, se retiene allí. Se pueden observar dos o más tipos de ligandos unidos a receptores, como LDL y transferrina, en la misma cavidad o vesícula revestida, se piensa que la polimerización regulada de la clatrina hace que las cavidades se expandan y eventualmente formen vesículas revestidas con clatrina.18
Exocitosis y endocitosis[editar]_Endocitosis_y_b)_exocitosis.jpg)
La exocitosis de vesículas y la endocitosis son eventos biológicos fundamentales que presentan cierta relación, pues la liberación de neurotransmisores en la sinapsis se basa en un ciclo sostenible entre estos dos procesos.19 La vesícula de exocitosis, haciendo uso de la membrana plasmática, libera contenido vesiculares para ejercer diversas e importantes funciones. Entre ellas están: la participación en la secreción de los transmisores de las neuronas, proceso esencial para las funciones cerebrales; la secreción neuronal de péptidos(como el neuropéptido Y) y hormonas (por ejemplo, la vasopresina, oxitocina), brinda una regulación del trabajo y del estado mental; la secreción de insulina de las células pancreáticas para regular el nivel de glucosa en la sangre; la secreción de catecolaminas y péptidos (implicado en la respuesta al estrés); además, la exocitosis desempeñada por las células sanguíneas para las respuestas inmunitarias.
Las funciones de la exocitosis en el organismo son de vital importancia. El papel de la endocitosis también adquiere gran relevancia puesto a que tras la exocitosis continua, se requiere de la endocitosis para mantener la estructura de la terminal del nervio, además de asegurar la disponibilidad funcional de las vesículas sinápticas. La membrana vesicular y las proteínas se recuperan a partir de la membrana plasmática por medio de este proceso.1920 Para este fin la endocitosis realiza el reciclaje de vesículas y protege a las células secretoras de la hinchazón o contracción.
Se proponen dos modelos para el funcionamiento de este procedimiento, el primero sugiere que las vesículas se someten a una secuencia reversible de exo-endocitosis, por medio del cual se preserva la identidad bioquímica de las vesículas tanto durante como después de la conexión transitoria entre la membrana de la vesícula y la membrana plasmática. El otro modelo establece que durante la exocitosis, la membrana de la vesícula se añade totalmente a la membrana plasmática y la recuperación se da tiempo después en un punto diferente de la membrana plasmática por endocitosis mediada por clatrina.
La incorporación de sustancias externas por parte de las células animales es esencial para su supervivencia. Ya vimos que algunas moléculas salvan la barrera que supone la membrana plasmática mediante difusión directa, a través de canales, mediante transportadores o por bombas (ver Transporte de membrana). Sin embargo, hay una manera de incorporar grandes cantidades de moléculas al interior de la célula de una sola vez: endocitosis o incorporación de moléculas englobadas por membranas, sobre todo vesículas. De la misma manera que hay un viaje de ida y fusión de vesículas con la membrana plasmática existe un proceso de formación de vesículas en la membrana plasmática, las cuales se fusionan posteriormente con compartimentos internos, principalmente con los endosomas.
Por tanto, una de las funciones de la endocitosis es la incorporación de moléculas externas en grandes cantidades, principalmente para su degradación. Pero también sirve para reciclar moléculas de la membrana plasmática que formarán parte de las propias vesículas o compartimentos que se formen. Otra función menos aparente es compensar los procesos de exocitosis, es decir, eliminar el exceso de membrana plasmática añadida por las vesículas de exocitosis y mantener así una superficie de membrana estable y funcional.

Las moléculas pueden ser incorporadas por endocitosis de forma específica unidas a receptores de la membrana plasmática o de manera inespecífica en disolución o pinocitosis.
En la incorporación masiva, las moléculas extracelulares pueden entrar al interior de la vesícula de forma inespecífica, en solución, o de forma específica unidas a receptores de membrana. El término pinocitosis se refiere a este tipo de endocitosis inespecífica de moléculas disueltas. Sin embargo, en la mayoría de las rutas de endocitosis existe un proceso de incorporación de moléculas de manera específica, bien por la acción de receptores de membrana o por su asociación con ciertos dominios lipídicos de membrana que favorecen la atracción de determinadas moléculas. No cabe duda de que parte del contenido de cualquier vesícula que se forme en la membrana plasmática tendrá moléculas disueltas que se hayan colado en el interior de la vesícula de manera inespecífica. Por tanto, en mayor o menor medida todas las rutas de endocitosis realizan pinocitosis. Este material en disolución es mayoritario en la macropinocitosis, donde la incorporación inespecífica de moléculas es su principal característica, como veremos más adelante. Hay que tener en cuenta que durante cualquier tipo de endocitosis también se incorporan lípidos y proteínas de la membrana plasmática, que son las que forman la membrana de la propia vesícula.
Al mecanismo de incorporación de moléculas específicas reconocidas por receptores de la membrana plasmática se le llama endocitosis mediada por receptor. Se han descrito unos 25 tipos de receptores que actúan en este tipo de endocitosis. Con ellos la célula puede incorporar de forma muy eficiente moléculas o partículas que se encuentran disueltas a bajas concentraciones. Estas moléculas se unen a sus receptores y los complejos receptor-ligando convergen en una zona de la membrana plasmática donde se produce la formación de la vesícula que posteriormente viaja hacia el interior celular. El ejemplo más llamativo es la captación de colesterol por parte de las células, el cual se transporta en la sangre unido a proteínas como, por ejemplo, las lipoproteínas de baja densidad (LDL). Las LDL son unos complejos que contienen una gran cantidad de moléculas de colesterol rodeadas por una monocapa lipídica y poseen una molécula proteica que sobresale al exterior. Cuando una célula necesita colesterol sintetiza receptores para los LDL y los traslada a la membrana plasmática. Entonces se produce el reconocimiento entre receptor y LDL, ambos se unen y se agrupan en una zona de la membrana plasmática donde se produce una invaginación. Una vez formada la vesícula, se dirige a orgánulos intracelulares donde las LDL son digeridas y el colesterol es liberado y metabolizado. Cuando se produce algún impedimento en la captación de colesterol, fundamentalmente por fallos en el reconocimiento por parte de los receptores de LDL o por su ausencia, el colesterol se acumula y puede producir arterioesclerosis e infarto de miocardio.

Distintos tipos de endocitosis.
Se han descrito diversos tipos de endocitosisdependiendo del tamaño de la vesícula, la naturaleza del material a incorporar y del mecanismo de formación de la vesícula, pero nosotros los vamos a agrupar en: endocitosis mediada por vesículas recubiertas de clatrina, endocitosis mediada por caveolas, endocitosis mediada por vesículas no recubiertas y macropinocitosis. Vamos a estudiar también a la fagocitosis, una endocitosis un tanto especial porque es un proceso de incorporación de grandes partículas como bacterias o restos celulares, también englobados en membranas.
Vesículas recubiertas de clatrina. Es el principal mecanismo por el que se incorporan proteínas integrales y lípidos de la membrana plasmática, así como macromoléculas extracelulares que generalmente no exceden los 156 nm, incluyendo algunos virus. Las vesículas se forman en áreas de la membrana plasmática donde se encuentra la proteína clatrina, que es citosólica. En los fibroblastos estas áreas suponen un 2 % del total de la superficie de la membrana plasmática. La clatrina posee una estructura con tres brazos que se ensamblan entre sí formando pentágonos. Su estructura y su manera de asociarse parece que ayudan a la invaginación y cierre de la vesícula. La polimerización de la clatrina forma vesículas de unos 120 nm. Entre la clatrina y la membrana celular se disponen otras proteínas denominadas adaptadoras que ayuda al ensamblaje de las moléculas de clatrina para formar una especie de cesta que engloba a la vesícula. Las proteínas adaptadoras son las que realmente van a decidir qué tipo de receptores de la membrana plasmática, junto con sus ligandos, van a formar parte de las vesículas, puesto que interaccionan con el dominio citosólico de las proteínas integrales. Una vez que la vesícula se ha cerrado e internalizado, la clatrina de desensambla y la vesícula puede ir a orgánulos específicos dentro de la célula, normalmente endosomas tempranos.
 Formación y fusión de vesículas.
Caveolas. Se describieron en los años 50 del siglo XX por P. Palade gracias a imágenes de microscopía electrónica. Son unos pequeñas invaginaciones en la membrana plasmática (45-80 nm) presentes en la mayoría de las células eucariotas que posteriormente se transforman en vesículas, aunque la proporción en que esto ocurre no está del todo clara. Son especialmente abundantes en las células endoteliales, musculares y adipocitos. Su membrana se caracteriza por poseer una proteína llamada caveolina, además de proteínas periféricas ancladas a glicosilfosfatidil-inositoles, esfingolípidos (esfingomielina y glicoesfingolípidos) y colesterol. La propia existencia de caveolina hace que las células formen caveolas. Hay de 100 a 200 moléculas de caveolina por caveola y existen diferentes tipos en una sola caveola. La caveolina 1 se expresa en el músculo liso y en la mayoría de las células no musculares, y es esencial para la formación de las caveolas en estas células. La caveolina 2, que se expresa junto con la caveolina 1, no es necesaria para la formación de las caveolas. La caveolina 3 se expresa en el músculo estriado, cardiaco y algunas otras células no musculares. Es esencial para formar caveolas en estas células. Las cavinas son otras proteínas presentes típicamente en las caveolas. No sólo se forman caveolas en la membrana plasmática sino que también se observan en el aparato de Golgi. Así que podría funcionar como mecanismo de transporte entre el aparato de Golgi y la membrana plasmática para determinados tipos de moléculas. Cuando se internan desde la membrana plasmática, las vesículas resultantes de las caveolas se fusionan con los endosomas tempranos, aunque algunos autores consideran que lo hacen con un tipo especial de endosoma denominado caveosoma.
Las funciones de las caveolas, sin embargo, no están del todo claras. Aunque se ha propuesto un papel relevante en la endocitosis, esto no parece tener un gran soporte experimental. El papel de las caveolas como transportador de moléculas en la endocitosis general de una célula no parece ser muy importante, al menos si excluimos células endoteliales, musculares y adipocitos, puesto que sólo el 5 % de las caveolas se convierten en vesículas, al menos en células en cultivo. Se han propuesto otras funciones que incluyen modulación de la transducción de señales celulares puesto que contienen numerosos receptores en sus membranas que son eliminados de la superficie celular como los receptores tirosina quinasa, el tráfico de lípidos entre la membrana y orgánulos interno, incluso se han postulado como participantes en los mecanismos de supresión de algunos tumores. Moléculas como la toxina colérica, el ácido fólico y otras moléculas entran a la célula gracias principalmente a las caveolas. También se ha sugerido que la variación en el número de caveolas de una célula podría influir en otros tipos de endocitosis como los mediados por clatrina, gracias al secuestro de moléculas en las propias caveolas y el consiguiente cambio de composición de la membrana plasmática. Una teoría que está recibiendo más apoyo experimental es su papel en la regulación de la tensión mecánica de las membranas, de manera que la presencia de más o menos caveolas contrarrestaría tensiones en la membrana de la células. Esto explicaría por qué es abundante en endotelios, células musculares y adipocitos, cuyas membranas estarían sometidas a cambios bruscos de tensión.
Endocitosis de vesículas que no dependen de la clatrina ni de la caveolina. Estas vías se han descubierto porque se siguen produciendo procesos de endocitosis cuando se bloquean selectivamente las vías mediadas por clatrina y por caveolina. Algunas toxinas como las del cólera entran preferentemente por esta vía. No se conoce en detalle cuales son los mecanismos por los que este tipo de endocitosis selecciona a las moléculas que transportan. Se sospecha que se pueden concentrar e incorporar moléculas gracias a la acción de las balsas de lípidos. Tampoco se conoce si la formación de estas vesículas es un proceso regulado o no.
Macropinocitosis. Es un proceso mediante el cual se incorporan grandes cantidades de fluido extracelular. En la superficie celular se crean evaginaciones a modo de ola cuyo frente cae sobre la membrana plasmática y se fusiona con ella formando una gran vesícula interna o macropinosoma. El mecanismo de formación de los macropinosomas involucra a los mismos componentes que actúan durante la fagocitosis: los filamentos de actina y las proteínas motoras miosina. La macropinocitosis no sólo se utiliza para captar alimento, como ocurre en las amebas, sino que también sirve para renovar la membrana plasmática, se activa durante el movimiento celular para transportar grandes porciones de membrana hacia el frente de avance, incluso algunas bacterias son capaces de inducirla para introducirse en los macropinosomas y así evitar la fagocitosis.
Fagocitosis. Es un tipo especial de endocitosis que consiste en la incorporación de partículas de gran tamaño como son bacterias, restos celulares o virus. Este mecanismo lo llevan a cabo células especializadas como son los macrófagos, neutrófilos y las células dendríticas. Un ejemplo claro son los macrófagos que fagocitan a los complejos formados por inmunoglobulinas unidas a otras partículas que pueden ser virus o bacterias. También son los encargados de eliminar miles de glóbulos rojos al día. Los protozoos utilizan este mecanismo para alimentarse. El proceso de fagocitosis supone un reconocimiento de la partícula por parte de la célula mediante receptores de membrana y la emisión de unas protuberancias laminares o pseudópodos de citoplasma rodeados por membrana. Este proceso está mediado por los filamentos de actina y las proteínas motoras miosina. Tales protuberancias rodean a la partícula, fusionan sus frentes de avance y encierran a la partícula formando una gran vesícula o fagosoma que se separa de la superficie y se interna en la célula para ser digerida. La fagocitocis requiere de una señal de reconocimiento para disparar el proceso. Una vez formado el fagosoma se fusionará con los lisosomas para la degradación de su contenido.
|





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
No hay comentarios:
Publicar un comentario