Un ojo compuesto es un órgano visual que se encuentra en ciertos artrópodos como insectos y crustáceos. Consiste en la agrupación de entre 12 y varios miles (6.300 en Apis mellifera) de unidades receptivas. Estos tipos de ojos no tienen una lente central, lo cual implica una baja resolución de imagen. Asimismo, los ojos compuestos son capaces de detectar movimientos rápidos, ven un amplio rango de ángulos sólidos y en algunos casos, perciben la polarización de la luz.1
Los ojos compuestos se encuentran principalmente en los insectos, y están formados por muchas facetas simples llamadas omatidios que dan una imagen en mosaico, no imágenes múltiples, como a menudo se cree.2 Cada omatidio consiste en una lente y un rabdoma, el cual está compuesto de grupos de células receptoras visuales (rabdomeros) puestas en paralelo o ligeramente giradas.
En insectos y crustáceos el ojo compuesto de los adultos se desarrolla en una sección de la cabeza diferente de la región en que las larvas tiene su ojo medio. Los nuevos omatidios se van añadiendo en hileras semicirculares en la parte de atrás del ojo durante el primer paso del desarrollo del ojo. Esto hace que los omatidos sean cuadrados, pero después, durante el desarrollo se vuelven hexagonales.

Anatomía del ojo compuesto de una mosca. En sentido horario: células pigmentadas primarias, lentes, conos cristalinos, células retinales, rabdomas, células pigmentadas secundarias, axones
Fisiología del ojo compuesto
Percepción del movimiento
El ojo compuesto es excelente en la percepción de movimiento. Si un objeto cruza el campo visual, los omatidios se encienden o apagan alternativamente. Por este "effecto de parpadeo" los insectos responden mejor a objetos en movimiento que a los que permanecen estacionarios. Las abejas, por ejemplo, visitan con preferencia flores movidas por el viento.4
Resolución óptica y sensibilidad
La agudeza visual es la capacidad del sistema de visión para percibir, detectar o identificar objetos especiales con unas condiciones de iluminación buenas.5 En el ojo compuesto la resolución o agudeza visual depende del número de omatidios y de la distancia entre ellos. Esto no permite reducir el tamaño de los omatidos para aumentar su número e incrementar la agudeza visual. Por lo tanto los ojos compuestos nunca alcanzan el grado de resolución de los ojos de vertebrados.6
Aquellos artrópodos activos cuando la luz es baja (cangrejo de río, mantis) desplazan los pigmentos de los omatidios hacia la base de las células pigmentadas. Esto permite que la luz entre a cada omatidio en un ángulo más ancho y que pase a los omatidios adyacentes estimulando también éstos en consecuencia. Así, la imagen es más pobre pero la percepción del nivel de luz aumenta. Probablemente el mantis sólo puede distinguir la ausencia o presencia de luz en condiciones de penumbra.
Visión del color
Muchos insectos perciben colores. Esto requiere dos o más pigmentos visuales, cada uno capaz de absorber luz de diferente longitud de onda.
En la abeja doméstica cuatro de las células visuales en cada omatidio responde optimamente a la luz amarillo-verdosa (544 nm); dos responden a la luz azul (344 nm) y las dos restantes responden a la luz ultravioleta (344 nm).
Este sistema permite que las abejas distingan los colores incluyendo luz ultravioleta, pero no el rojo. Los estudios comportamentales confirman esto. La visión ultravioleta de las abejas les sirve para encontrar polen y néctar en ciertas flores que tienen guías de néctar que estos insectos pueden ver aunque sean invisibles para los humanos.
Las mariposas monarca usan luz ultravioleta para orientación en su larga migración de más de 4.000 Km.4
Hay dos tipos básicos de ojos compuestos
- El ojo de aposición que se puede dividir en dos grupos. El típico ojo de aposición tiene una lente que enfoca la luz proveniente de una dirección sobre el rabdómero, mientras que la luz proveniente de otras direcciones se absorbe en las paredes oscuras del omatidio. El otro tipo de ojo de aposición se encuentra en el orden Strepsiptera, en el cual cada lente forma una imagen, y las imágenes se combinan en el cerebro. Este ojo se llama ojo de superposición neuronal u ojo esquizocroal compuesto.
- El segundo tipo se llama ojo de superposición. Se divide en tres tipos: superposición refractante, superposición reflectante y superposición parabólica. El ojo de superposición refractante tiene una abertura entre la lente y el rabdómero y no tiene pared. Cada lente refleja la luz en un ángulo igual al ángulo que la recibe. El resultado es la formación de la imagen en la mitad del radio del ojo, donde hay situadas las testas de los rabdómero. Este tipo de ojo se encuentra normalmente en insectos nocturnos. En los ojos compuestos de superposición parabólica, que se encuentran en artrópodos como las efímeras, cada faceta de la superficie del ojo contiene una superficie parabólica que recibe la luz de un reflector y la enfoca sobre una matriz de sensores. Los crustáceos de cuerpo largo como gambas, langostinos y langostas son los únicos que tienen ojos de superposición reflectante, que también tienen una obertura pero que en lugar de lentes utilizan diedros de espejos.
Hay algunas excepciones de los casos anteriores. Algunos insectos tienen lo que se denomina un ojo compuesto de lente simple, que es un caso intermedio entre el ojo compuesto de superposición y el ojo de lente simple que se encuentra en los animales de ojos simples.
El cuerpo del Ophiocoma wendtii, un tipo de ofiuroideo, está cubierta de omatidios, convirtiendo toda su piel en un ojo compuesto.

Los ojos esquizocroales son un tipo de ojos compuestos formados por varias lentes separadas por un material llamado esclerótida.1 Este tipo de ojos es exclusivo de un orden de trilobites, los Phacopina.2 Parece ser que este tipo de ojos serían útiles en condiciones de poca luminosidad, por lo que es probable que los Phacopina fueran animales de hábitos nocturnos.1 Otros análisis sugieren la posibilidad de que estos ojos permitieran una visión estereoscópica de 360º.1 Con la extinción de estos trilobites en el Devónico superior desaparecieron los ojos esquizocroales, conservándose solamente como fósiles.
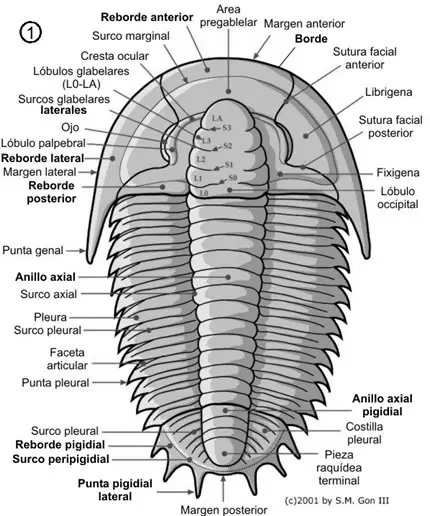
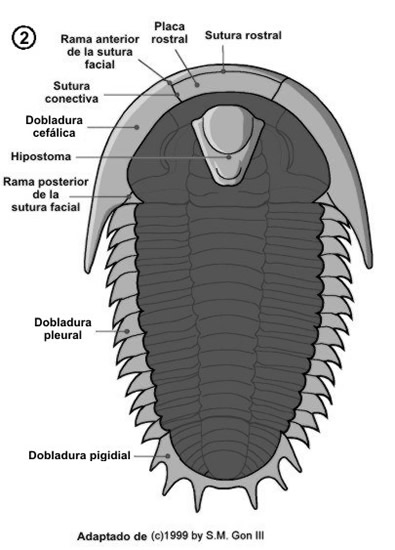
Este esqueleto se encuentra atravesado por numerosas sedas con función sensitiva y presenta numerosas ornamentaciones crateriformes o tuberculosas.
Región cefálica o céfalon
En el céfalon la parte que más destaca es la glabela, que no es más que la parte anterior del lóbulo axial, convexa y que presenta un número variable de surcos transglabelares o surcos laterales que corresponden al número de segmentos que forman esta región. En todos los trilobites el céfalon es una placa formada por un número variable de segmentos fusionados en los que se encuentran los ojos, las suturas faciales y el hipostoma, y la gran variedad de morfologías que puede adoptar son en la gran mayoría de las especies caracteres identificativos.
En el esqueleto externo del céfalon se pueden diferenciar un cranidio, formado por la glabela y las mejillas fijas o fixígenas; lateralmente al cranidio están las librígenas o mejillas libres, separadas del mismo por la sutura facial de la que posteriormente trataremos (ver fig. 3). Las mejillas corresponden a las regiones pleurales cefálicas.
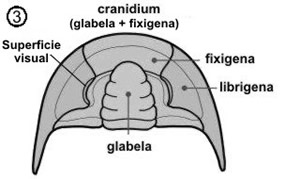
En el céfalon se dividen zonas atendiendo a las suturas, placas y posición que ocupan de gran importancia en la determinación de las diferentes especies (ver fig. 4).
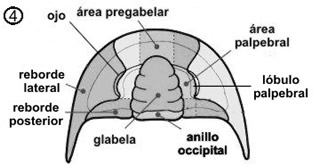
La glabela varía considerablemente entre las especies, tanto en forma como en estructura, siendo en unos casos bastante pronunciada con lóbulos laterales o en otros menos conspicua. Se cree que el estómago se encontraba entre ella y el hipostoma de la parte inferior, por lo que su tamaño guardaría una estrecha relación con el mismo y por consiguiente con la dieta de la especie. Variaciones en su tamaño y el número de estrías pueden considerarse indicadores del ambiente y dieta.
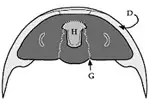
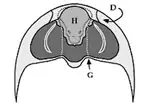
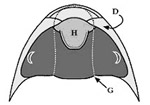
El hipostoma (fig. 2), situado en la zona ventral de la región cefálica, es una pieza de exoesqueleto que se cree que formaba parte de la boca. Su borde anterior se encuentra bajo el borde anterior de la glabela por lo general. Puede adoptar gran variación morfológica usada como caracter identificatorio y básicamente se diferencian 3 tipos:
- Flotante: se encuentra separado y libre del pliegue anterior y su borde anterior coincide con el de la glabela.
- Conectado: se encuentra fijo al pliegue anterior de la placa rostral y su borde coincide con el anterior de la glabela.
- Desplazado: semejante al anterior, pero el borde anterior no coincide con el de la glablela.
Flotante
Conectado
Desplazado
El hipostoma flotante muestra diseños bastante conservadores a lo largo de su evolución; su forma suele ser ovoide y carente de extensiones u ornamentaciones. La alimentación generalista a base de partículas es la causa de las formas tan simple y poco variadas de su hipostoma.
Por el contrario, los otros dos modelos han sufrido una mayor diversificación que originó una serie de modelos muy variados en forma y tamaño que ha servido para clasificar muchas especies de trilobites y especular sobre sus hábitos alimentarios. Se encuentran en especies que se cree que fueron depredadoras, por lo que su anclaje a la placa rostral proporcionaría más solidez al mismo y ante los posibles ataques a las presas; la especialización de formas estaría relacionada con diferentes tipos de presas o comportamientos alimenticios.
Las suturas están relacionadas con las mudas. No hemos de olvidar que los trilobites son artrópodos y como tales tienen un esqueleto externo; el crecimiento del animal viene entonces marcado por las mudas de este exoesqueleto, que debe romper por unas líneas fijas (las suturas) para salir con un nuevo exoesqueleto de mayor tamaño. Estas suturas indican el lugar de la región cefálica por donde se separan las placas para permitir la salida del animal mudado y por lo general discurren desde el borde anterior del céfalon, alcanzan la superficie visual, separando la superficie visual el lóbulo palpebral, y continúan hacia el borde posterior de la misma. Hay tres tipos principales de suturas faciales en función del lugar donde finalizan en la zona posterior (fig. 5):
- Propárica: el final se encuentra en el borde anterior al ángulo o espina genal.
- Gonatopárica: el final de la sutura desemboca posteriormente al ángulo genal o muy próximo a él.
: el final se encuentra en cualquier punto del borde posterior cefálico. En alguno grupos muy concretos el recorrido de esta sutura corre junto al margen posterior, llamándose entonces marginopárica.

La sutura facial además parte la zona dorsal de la mejilla en las dos que vimos anteriormente, la fixigena y la librigena; la segunda, la mejilla libre, se desarticula durante la muda y también frecuentemente tras la muerte del animal, por lo que es fácil encontrar fósiles sin esta pieza.
Las suturas dorsales no finalizan en los margenes laterales, sino que se extienden por la región ventral de la zona cefálica a lo largo de la dobladura cefálica, la placa rostral y el hipostoma.
En el céfalon también se encuentran los ojos. Los trilobites fueron los primeros seres vivos en desarrollar un sistema visual y éste es bastante semejante al que tienen los crustáceos o insectos actuales, los ojos compuestos, formados por múltiples facetas independientes cuyos campos visuales individuales procesados por el cerebro forman la imagen completa. Fueron además los primeros en desarrollar este tipo de ojo. Se encuentran el borde externo de la fixigena, a ambos lados de la glabela y adyacentes a la sutura facial. Hay grupos de trilobites que carecen de ojos (Agnostina), mientras que otros los pierden tras iniciar una reducción de su tamaño de forma secundaria y otros sufren el proceso contrario, desarrollando ojos de gran tamaño en función del ambiente en el que evolucionaron.
En los vertebrados es posible que la lente ocular o cristalino varíe su forma para lograr enfocar; sin embargo, en los trilobites y demás artrópodos las lentes son rígidas, por lo que no pueden modificarse para enfocar. Para conseguirlo disponen de una segunda lente que combinada con la anteiror soluciona el problema del foco.
Hay tres tipos de ojos en los trilobites:
- Holocroal: presente en la mayor parte de los órdenes de trilobites, están formados por hasta 15000 facetas pequeñas. La membrana de la córnea cubre sólo la superficie de las facetas que están en contacto las unas con las otras.
- Esquizocroal: este tipo se considera una innovación presente en el grupo de los Phacopida. No tiene tantas facetas como el grupo anterior, solamente alrededor de unas 700, pero su tamaño es mayor. Cada una de las facetas tiene una córnea individual y se encuentran separadas por una capa esclerotizada relativamente gruesa; la membraba de la córnea penetra en la parte interna de la faceta y se llama en esta zona membrana intrascleral.
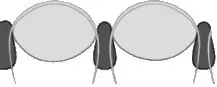
- Abatocroal: solamente se han encontrado en el género Eodiscina del Cámbrico. Sólo tiene unas 70 lentes o facetas de pequeño tamaño, cada una con una córnea individual y separada del resto de facetas por una capa esclerotizada no tan gruesa como la presente en el modelo ocular anterior. La córnea no penetra en el interior de la faceta.

Estos ojos se encuentran en número par y en posición dorsal, generalmente vertical para lograr una visión completa del entorno, en muchos casos de 360º. Muchos trilobites presentan además dos ojos simples u ocelos conocidos como máculas, en posición ventral y en el hipostoma, que podrían semejar a los ocelos de las larvas nauplius de los artrópodos y que no desaparecerían durante la ontogenia, al igual que ocurre en otros muchos artrópodos actuales.
Entre los ojos de los trilobites existe una gran variedad morfológica que responde a las diferentes adaptaciones que mostraban a los diferentes nichos en los que se encontraban. Muchas de las especies más primitivas tenían ojos con forma de media luna; el aumento de estos hasta adoptar una forma casi cónica en sección (ojos esquizocroales) permitió aumentar el campo visual, algo que en géneros nadadores como Opipeuterella hizo que sus enormes ojos le proporcionaran una visión de 360º. Especies bentónicas como las del género Cryptolithus tenían los ojos bastante reducidos o carecían de ellos, y otras que vivían en zonas con detritos o vegetación podían mostrar los ojos levantados del cuerpo sobre pedúnculos para lograr una mejor visión de los alrededores como ocurre en Neoasaphus. Especies de aguas profundas, donde no había luz, los ojos no les proporcionaban ninguna ventaja por lo que fueron reduciéndose secundariamente hasta poder acabar por desaparecer.
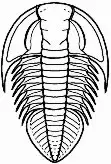
Polypleuraspis
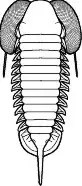
Opipeuterella
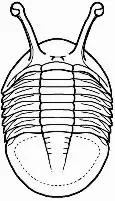 Neoasaphus
Neoasaphus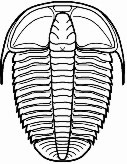
Polypleuraspis
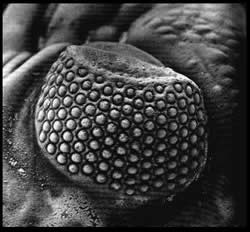
Ojo esquizocélico de Phacops
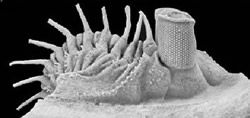
Erbenochile erbeni, encontrado en el Devónico de Marruecos en 2003, que muestra un extraordinario desarrollo ocular.
En el céfalon también se encuentran un par de antenas que surgen de los laterales del hipostoma y que en la mayoría de los casos se pierden durante la fosilización debido a su extrema fragilidad.
No hay comentarios:
Publicar un comentario